|
|
Жизнь |
|
|
|
| ||||
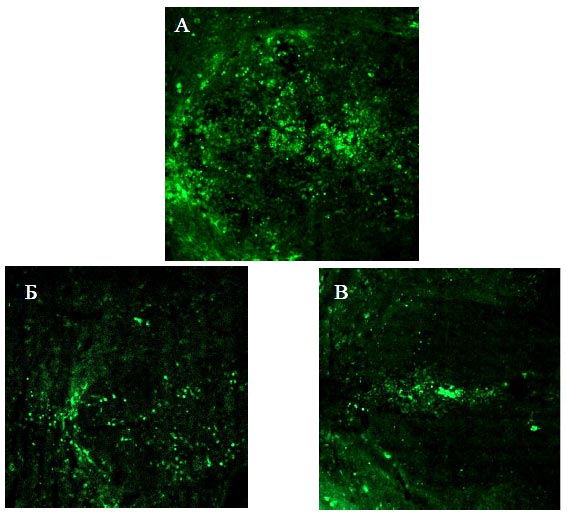
3.3. Эффективность посттравматической регенерации при прямой генной терапии плазмидой pBud-VEGF-FGF2Трансфекционная активностьИнъекция плазмиды с геном EGFP подтвердила, что введенная в область травмы плазмида сохраняет активность и трансфицирует клетки спинного мозга. На 14-е сутки после КТСМ выявлены скопления интенсивно светящихся клеток на расстоянии 0,2 и 0,5 см от эпицентра травмы в каудальном и ростральном направлениях (Рис. 58).
Рис. 58. Свечение EGFP на 14 сутки после КТСМ и инъекции в зону повреждения плазмиды pBud-EGFP (контроль) на расстоянии: А - 0,2 см в каудальном, Б - 0,5 см в ростральном, В - 0,5 см в каудальном направлении от эпицентра травмы, дорсальные канатики. Флюоресцентная микроскопия. Увеличение 10х10 Восстановление функцииОднократная инъекция в зону повреждения спинного мозга крысы плазмиды, экспрессирующей гены vegf и fgf2, стимулирует восстановление двигательной активности задних конечностей. В первые 6 суток эксперимента после КТСМ двигательная активность задних конечностей, как в опытной группе животных, так и в контрольной, отсутствует. После прохождения фазы спинального шока начинается период относительно быстрого нарастания двигательной активности (Рис. 59), после чего уровень функциональности стабилизируется (фаза плато).
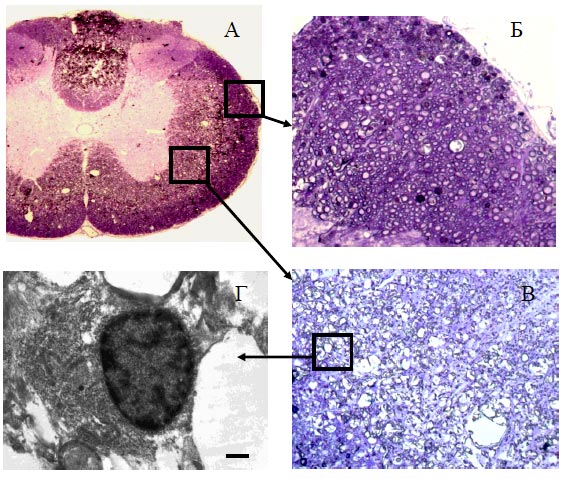
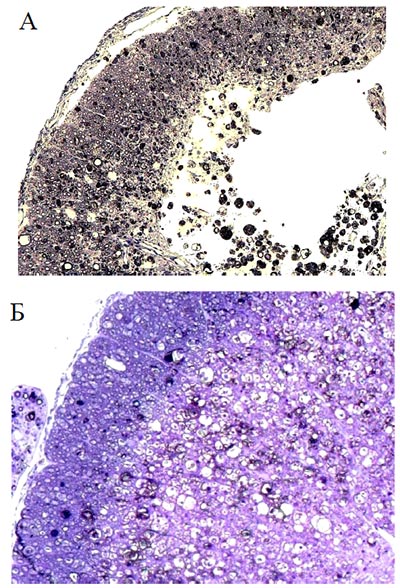
Рис. 59. Результаты теста «ВВВ» при введении плазмиды pBud-VEGF-FGF2. По оси ординат - баллы, по оси абсцисс - сутки после КТСМ. Черные квадратики - опыт, белые кружочки - контроль (pEGFP) Дальнейшее тестирование животных в открытом поле выявило более выраженное восстановление двигательной активности при инъекции плазмиды pBud-VEGF-FGF2: с 3,5 баллов на 7-е сутки после КТСМ до 14,5 баллов к концу эксперимента. Средний прирост показателя «ВВВ» к 30-м суткам в опыте (pBud-VEGF-FGF2) составил 11 баллов, а в контроле (pEGFP) - 4 балла (р<0,05). Приведенные на рис. 59 зависимости имеют уже отмечавшийся нами ранее двухфазный вид. Сами по себе темпы восстановления двигательной активности при введении плазмиды с генами нейротрофических факторов и плазмиды, несущей гены нейтрального белка, примерно одинаковы. Более чем трехкратное превышение показателя к моменту выхода на плато в опыте по сравнению с контролем обеспечивается увеличением времени, в течение которого действует механизм восстановления двигательных функций до его исчерпания. Достигнутая в опыте с непосредственным введением плазмиды pBud-VEGF-FGF2 длительность активной фазы восстановления (11 дней) превышала также длительность активной фазы при клеточноопосредованном введении той же плазмиды (5 дней), что обеспечило в первом случае на 2 балла более высокий уровень восстановления. Сохранность тканиВ опытах с введением в зону травматического повреждения плазмиды pBud-VEGF-FGF2 на расстоянии 5 мм от эпицентра повреждения центральная полость отсутствует. На поперечных срезах спинного мозга четко видно серое и белое вещество. Белое вещество не однородно (Рис. 60 (А)). Белое вещество, располагающееся по периферии органа содержит большое количества миелиновых волокон (Рис. 60 (Б)). В белом веществе, прилегающем непосредственно к серому веществу, количество миелиновых волокон значительно меньше (Рис. 60 (В)). К 30 суткам после травмы спинного мозга крысы инъекция плазмиды pBud-VEGF-FGF2 в область повреждения приводит к уменьшению области деструкции серого и белого вещества как в ростральном, так и каудальном направлениях от эпицентра травмы. Так, в опытной группе животных площадь участка повреждения на расстоянии 3 мм от эпицентра травмы в среднем на 55%, а на расстоянии 5 мм в среднем на 80% меньше, чем при инъекции плазмиды с геном EGFP (Рис. 63).
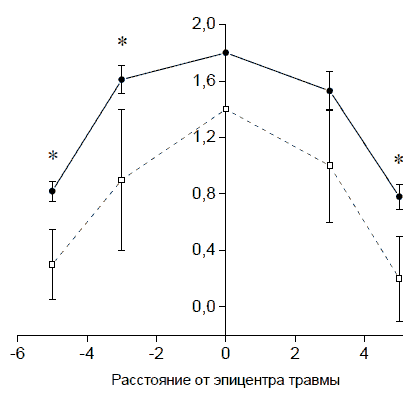
Рис. 60. Поперечный срез спинного мозга (А), фрагменты белого вещества (Б, В), и электроннограмма (Г) участка белого вещества на расстоянии 5 мм от эпицентра повреждения в ростральном направлении. Увеличение 10х10, 40х10, 8000. Длина отрезка 1 мкм При сравнении опытных групп, получавших генно-клеточную терапию (KKn+pBud-VEGF-FGF2) и прямое введение плазмиды pBud-VEGF-FGF2, площадь деструкции ткани в последнем случае оказалась в 1,5 - 2 раза меньшей (Рис. 61).
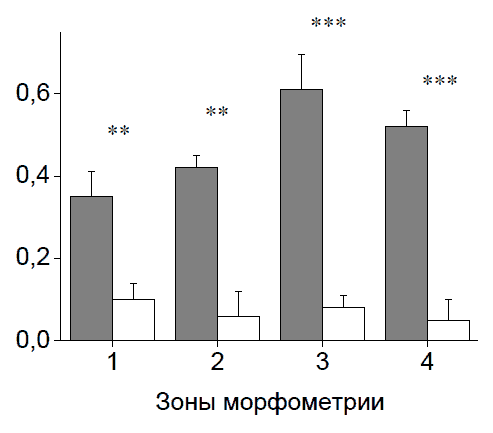
Рис. 61 . Площадь участка повреждения (в мм2 по оси ординат) на поперечных срезах спинного мозга. По оси абсцисс - расстояние в мм от эпицентра травмы: слева от оси ординат - в каудальном направлении, справа - в ростральном. Сплошная линия - KKn+pBud-VEGF-FGF2. Пунктирная линия - pBud-VEGF-FGF2. * р<0,05 Суммарная площадь патологических полостейИнъекция плазмиды pBud-VEGF-FGF2 в зону травматического повреждения к 30-м суткам эксперимента значительно уменьшает образование патологических полостей в спинном мозге по сравнению с контролем. На расстоянии 3 мм от эпицентра травмы во всех зонах морфометрии, как в ростральном, так и в каудальном направлениях, площадь патологических полостей в среднем в 5 раз меньше, чем в контроле (Рис. 62 и 63).
Рис. 62. Суммарная площадь патологических полостей (ось ординат в мм ) в контроле (pEGFP) и в опыте (плазмида pBud-VEGF-FGF2) на 30-е сутки после КТСМ на поперечном срезе спинного мозга на расстоянии 3 мм в каудальном направлении от эпицентра травмы. Серые столбики - контроль, белые столбики - опыт. По оси абсцисс - зоны морфометрии. ** р<0,005; *** р<0,001 Суммарная площадь патологических полостей в эксперименте с прямой генной терапией (pBud-VEGF-FGF2) на расстоянии 3 мм от эпицентра травмы в зонах 3 и 4 рострального фрагмента на 40% и 33%, а в зонах 2 и 4 каудального фрагмента - на 50% и 64% меньше, чем в эксперименте с клеточно-опосредованной терапией (ККП+pBud-VEGF- FGF2) (Рис. 62, 63, 64, 65).
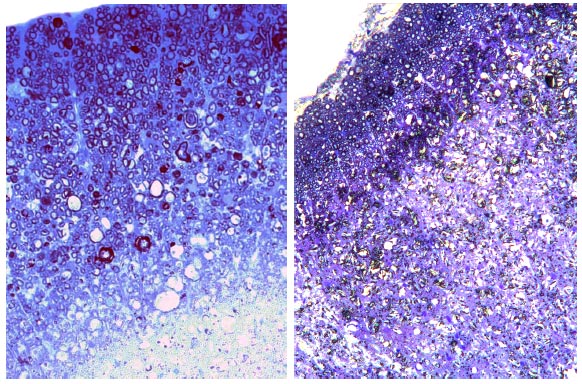
Рис. 63. Фрагмент белого вещества спинного мозга на поперечных срезах к 30м суткам на расстоянии 5 мм от эпицентра контузионного травматического повреждения на уровне Т8. А - контроль (pEGFP); Б - опыт (Bud-VEGF-FGF2). Окраска толуидиновым синим. Увеличение 40х10
Рис. 64. Суммарная площадь патологических полостей на расстоянии 3 мм от эпицентра травм зона 2: слева ККП+pBud-VEGF-FGF2, справа pBud-VEGF-FGF2. Фрагмент белого вещества спинного мозга. Срезы окрашены толуидиновым синим. Увеличение 40х10
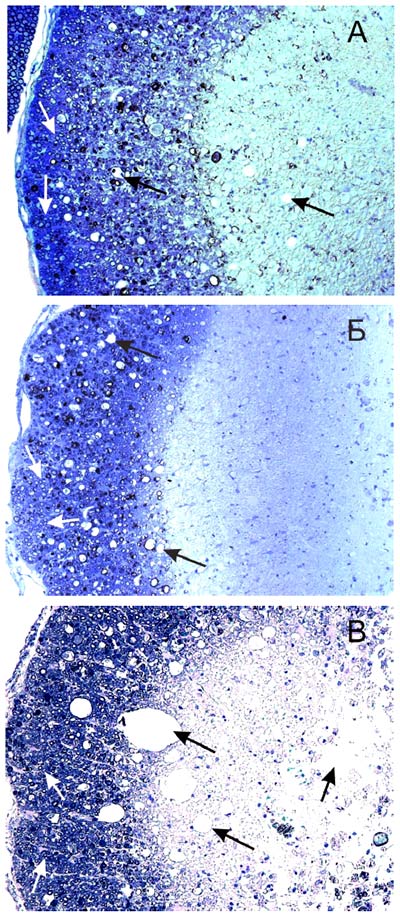
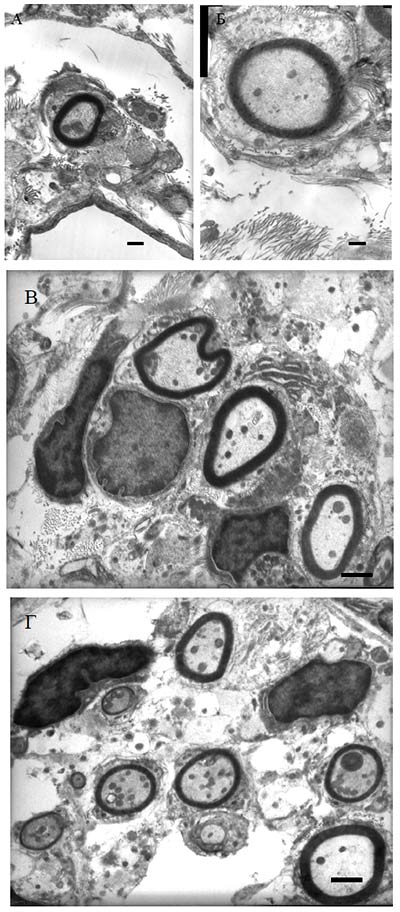
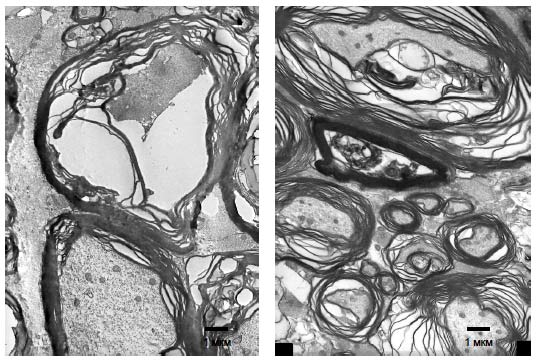
Рис. 65. Фрагмент белого вещества спинного мозга крысы (30 суток после травмы) на расстоянии 5 мм от эпицентра травы зона 3: А - при введении клеток крови пуповины, трасфицированных pBud-VEGF-FGF2, Б - при прямом введении плазмиды pBud-VEGF-FGF2, В - при введении клеток крови пуповины, транфицированных pEGFP. Черные стрелки указывают на полости, белые стрелки - на миелиновые волокна. Поперечные срезы окрашены толуидиновым синим. Увеличение 20х10 Количество миелиновых волоконНа 30-е сутки после контузионной травмы и инъекции в зону повреждения плазмиды pBud-VEGF-FGF2 количество миелиновых волокон на расстоянии 3 мм от эпицентра травмы достоверно не увеличивается по сравнению с контролем (pEGFP). Однако признаки деструкции большинства миелиновых волокон опытной группы животных менее выражены. При световой микроскопии миелиновые оболочки волокон в опыте имеют более четкие очертания и толщину, а сами волокна имеют больший диаметр, чем в контроле. При изучении волокон на электронно-микроскопическом уровне установлено, что миелиновые оболочки имеют незначительные локальные расслоения, характеризуются отсутствием вакуолей. Осевые цилиндры волокон в материале опытной группы животных менее деформированы. Они содержат микротрубочки, митохондрии и нейрофиламенты с более четкой структурой микротрубочек и митохондрий, чем в контроле (Рис. 65, 66 и 67).
Рис. 66 А-Г. Миелиновые волокна в белом веществе спинного мозга при введении в зону повреждения плазмиды pBud-VEGF-FGF2 (опыт). Эпицентр травмы. TEM. Увеличение 6000. Длина отрезков 500 нм
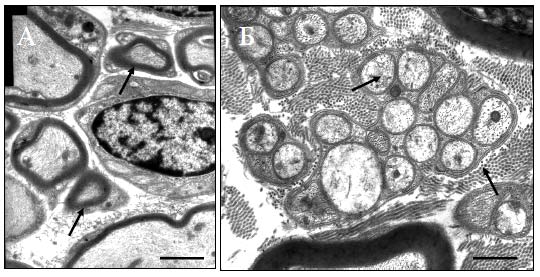
Рис. 67. Миелиновые волокна в белом веществе спинного мозга при введении в зону повреждения pEGFP (контроль) на расстоянии 5 мм от эпицентра травмы TEM. Увеличение 5000 На расстоянии 5 мм от эпицентра травмы в наружной части белого вещества в опыте наряду с волокнами обычной формы (Рис. 68, 69) обнаруживаются волокна с компактной миелиновой оболочкой диаметром меньше 2 мкм (указаны стрелками на рисунке 68 (А)), с хорошо выраженной базальной мембраной и окруженые пучками коллагеновых волокон. Наличие базальной мембраны и внеклеточного коллагена является признаком миелина периферического типа (Berthold et al. 1978) и отличает миелин-формирующие клетки от миелина центрального типа, образуемого олигодендроцитами. Выявляемые ТЕМ миелинизированные волокна имеют более тонкий, чем в норме, слой миелина с присоединенной извне асимметрично расположенной цитоплазмой, в которой просматриваются ядро и другие органеллы. Указанные признаки характерны для миелинобразующих шванновских клеток (Jasmin et al. 2000, Plant et al. 2002). В непосредственной близости от миелиновых волокон прослеживается большое количество безмиелиновых волокон (указаны стрелками на рисунке 68 (Б)), что не характерно для контрольной группы животных. Безмиелиновые волокна объединены в пучки, окруженные общей мембраной шванновских клеток. Пространство между пучками безмиелиновых волокон заполнено коллагеновыми волокнами. Подобная структура также характерна для волокон периферических нервов (Grisolia et al. 2006). Таким образом, однократная инъекция плазмиды pBud-VEGF-FGF2 в область контузионной травмы спинного мозга крысы стимулирует прорастание и ремиелинизацию волокон, опосредованную мигрирующими шванновскими клетками.
Рис. 68. Миелиновые волокна (А), безмиелиновые волокна (Б) в белом веществе спинного мозга на 30 сутки эксперимента на расстоянии 5 мм от эпицентра травмы при прямом введении плазмиды pBud-VEGF-FGF2. Увеличение на рисунке А 7500, на рисунке Б - 15000. TEM. Длина отрезка на рисунке А 1 мкм, на рисунке Б - 500 нм
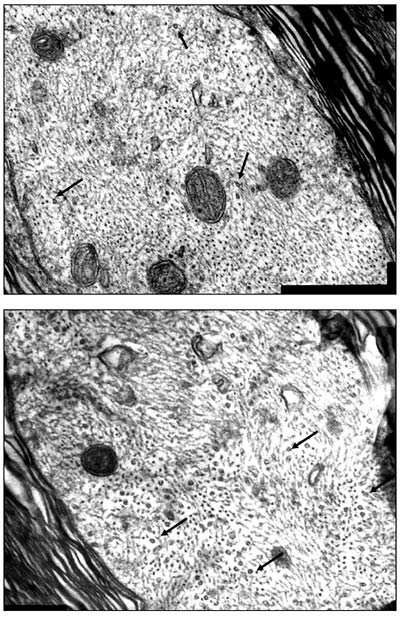
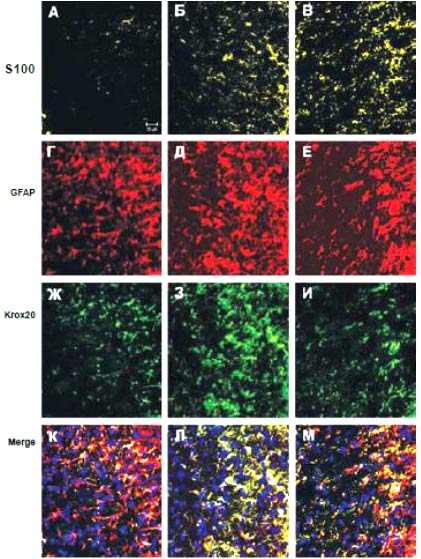
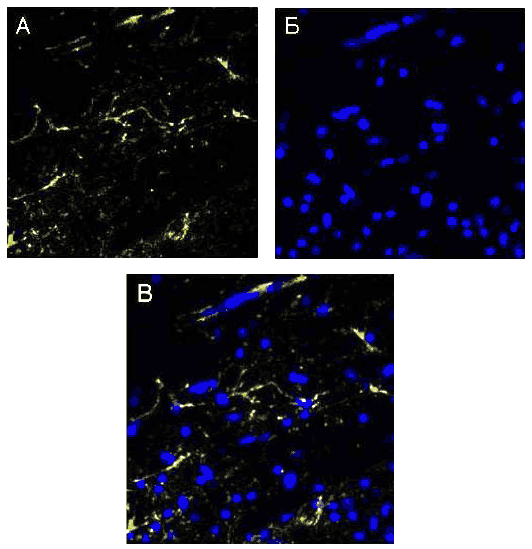
Рис 69. Ультраструктура миелиновых волокн от 2 мкм pBud-VEGF-FGF2. В осевых цилиндрах имеется много митохондрий и микротрубочек (на микротрубочки указывают стрелки). TEM. Увеличение 60000 Данные иммуногистохимииНа 30 сутки после КТСМ в группах животных ККП+pBud-VEGF-FGF2 и pBud-VEGF-FGF2 на расстоянии до 2 см в ростральном и каудальном направлениях от эпицентра повреждения обнаружены клетки, экспрессирующие специфические маркеры шванновских клеток (Рис. 70). Их количество и распределение в белом и сером веществе существенно различается в этих опытных группах от контрольной (КТСМ). При помощи непрямого иммунопероксидазного метода с выявлением Krox20 обнаружено увеличение количества Кгох20+-клеток во всех зонах морфометрии белого вещества на расстоянии 1 см в ростральном и каудальном направлении от эпицентра повреждения в обеих опытных группах по сравнению с контрольной (КТСМ) соответственно на 61% и 50%. К 30-м суткам после нанесения травмы во всех 3-х исследуемых группах в белом и сером веществе обнаружены клетки, одновременно экспрессирующие белки S100, GFAP и Krox20 (S100+/GFAP+/Krox20+- клетки). Для контрольной группы в области вхождения задних корешков характерно: отсутствие S100+-клеток и Кгох20+-клеток, преимущественно наличие GFAP+-клеток, небольшое скопление GFAP+/Krox20+-клеток и единичные S100+/GFAP+/Krox20+-клетки. Для обеих опытных групп в указанной области серого вещества характерно: наличие S100+-клеток и единичных Кгох20+-клеток, присутствие GFAP+-клеток, GFAP+/Krox20+-клеток и S100+/GFAP+/Krox20+-клеток. При этом в случае доставки генов нейротрофических факторов vegf и fgf2 на клеточных носителях (ККП+pBud-VEGF-FGF2) по сравнению с прямым введением плазмиды с терапевтическими генами (pBud-VEGF-FGF2) популяция S100+/GFAP+/Krox20+-клеток представляется наибольшей. В указанной области вхождения задних корешков в 1 опытной группе (ККП+pBud-VEGF-FGF2) особенно выражена популяция GFAP+/Krox20+- клеток, являющаяся также самой многочисленной.
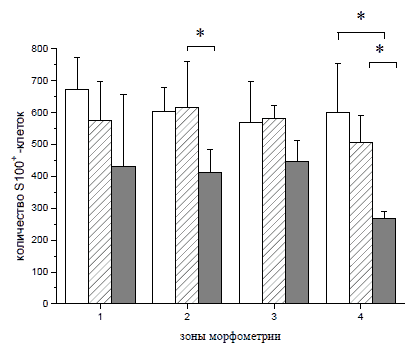
Рис. 70. Спинной мозг крысы. Экспрессия S100, GFAP и Krox20 в области вхождения задних корешков на расстоянии 1 см каудальнее эпицентра травмы: А, Г, Ж, К - контрольная группа животных; Б, Д, З, Л - 1 опытная группа с ведением ККП, трансфицированных плазмидой pBud-VEGF-FGF2; В, Е, И, М - 2 опытная группа с ведением плазмиды pBud-VEGF-FGF2; К, Л, М - объединение панелей по всем трем маркерам (Merge). Конфокальная микроскопия. Увеличение 40х10 Для 2 опытной группы (pBud-VEGF-FGF2) в области вхождения задних корешков характерно наличие большего количества S100+-клеток (Рис. 70 (В)) и GFAP+-клеток (Рис. 70 (Е)). В белом и сером веществе для всех экспериментальных групп характерно наличие единичных S100+-клеток округлой формы. Отмечены различия в имеющихся размерах тел и отростков S100+/GFAP+/Krox20+-клеток в белом и сером веществах, характерные для опытных и контрольной групп. Так, размер шванновских клеток, одновременно экспрессирующих белки S100, GFAP и транскрипционный фактор Krox20, в зонах белого вещества в 2 раза больше, чем в сером веществе и задних канатиках. К 30 суткам эксперимента количество S100+-клеток в наружных зонах белого вещества на расстоянии 1,5 см от эпицентра травмы в условиях прямой доставки генов возрастает на 46% (р<0,05) (Рис. 71). При введении клеток крови пуповины, трансфицированных плазмидой pBud-VEGF-FGF2, этот показатель увеличивается на 55% (р<0,05). При тройном иммунофлуоресцентом окрашивании с помощью антител против HSP25, GFAP и Krox20 в белом веществе выявлены клетки, экспрессирующие все три указанных маркера (HSP25+/GFAP+/Krox20+- клетки). В сером веществе и в области вхождения задних корешков данные клетки не обнаружены. Наибольшее количество HSP25+/GFAP+/Krox20+- клеток в области задних канатиков характерно для 1 опытной группы с доставкой генов vegf и fgf2 на клеточных носителях. Для 2 опытной группы с прямой доставкой плазмиды с теми же генами этот показатель снижен в 2 раза. В контрольной группе HSP25/GFAP/Krox20+-клетки в области задних канатиков отсутствуют, наблюдаются лишь единичные GFAP+/Krox20+- клетки и большое количество GFAP+-клеток. ШР25+-клетки в области вхождения задних корешков обнаружены лишь в 1 опытной группе (ККП+pBud-VEGF-FGF2).
Рис. 71. Количество S100-иммунопозитивных клеток в белом веществе спинного мозга крысы на расстоянии 1,5 см от эпицентра на 30 сутки эксперимента. Серые столбики - при трансплантации ККП, трансфицированных плазмидой pBud-VEGF-FGF2, белые столбики - при введении плазмиды pBud-VEGF-FGF2, темные столбики - контрольная группа с ведением ККП, трансфицированных плазмидой pEGFP. * - р<0,05 Полученные результаты свидетельствуют о том, что доставка генов vegf и fgf2 приводит к увеличению количества эндогенных шванновских клеток в области повреждения спинного мозга. Данный феномен можно объяснить либо увеличением срока выживания эндогенных шванновских клеток, либо повышением их способности к миграции в область повреждения и пролиферации. Эти данные согласуются с показанной ранее возможностью плазмиды с геном сосудистого эндотелиального фактора роста (vegf) усиливать пролиферацию шванновских клеток, увеличивать экспрессию в них белка vegf, тем самым улучшая способность к регенерации аксонов (Haninec et al. 2012). Способность усиливать пролиферацию шванновских клеток in vitro установлена и для фактора роста фибробластов 2 (fgf2) (Shen et al. 2005). Транскрипционный фактор Krox20 является надежным маркером миелинобразующих шванновских клеток и также экспрессируется клетками пограничной шапочки, которые активно мигрируют в область повреждения, где дифференцируются в шванновские клетки (Shen et al. 2005). Начало экспрессии Krox20 приурочено к стадии перехода предшественников в зрелые клетки (Topilko et al. 1994). Наличие белка S100 специфично как для астроцитарной глии, так и для незрелых шванновских клеток. Ранее GFAP считался высоко специфичным только для зрелых астроцитов, но исследования последних лет показали экспрессию данного белка в миелиннеобразующих шванновских клетках (Wang et al. 2010). Обнаружение S100+/GFAP+/Krox20+-клеток в нашем исследовании может свидетельствовать о наличии мигрирующих в спинной мозг шванновских клетках на стадии неполной дифференцировки в миелинобразующий клеточный тип. При этом среди всей популяции мигрировавших шванновских клеток встречаются и зрелые Кгох20+-клетки, способные к миелинизации аксонов. Увеличение количества Krox20+-клеток в 1 опытной группе (KKn+pBud-VEGF-FGF2) по сравнению с группой 2 (pBud-VEGF- FGF2) и контрольной (КТСМ) как в белом веществе, так и в области вхождения задних корешков говорит о стимулировании миграции и дифференцировке шванновских клеток в большей степени при доставке генов vegf и fgf2 при помощи ККП человека. Наиболее вероятно, что это связано со способностью к длительному выживанию трансплантированных клеток и поддержанию продукции ими терапевтических молекул. Способность к стимулированию миграции шванновских клеток в случае прямого введения плазмиды с генами vegf и fgf2 также имеет место, однако более низкие показатели по сравнению с 1 опытной группой (ККП+pBud- VEGF-FGF2), могут быть связаны с их низкой трансфекционной активностью при этом способе доставки или кратковременной продукцией нейротрофических факторов. Белок теплового шока HSP25 играет центральную роль в системе сигнализации, которая обеспечивает регенерацию нервных волокон (Murashov et al. 2001). Ранее показано, что HSP25 экспрессируется в глиальных клетках, в том числе реактивных астроцитах, находящихся в сером веществе (Pieri et al. 2001). По экспрессии белка теплового шока HSP25 в глиальных клетках можно судить о включении цитопротекторного механизма. Нами впервые обнаружена экспрессия белка теплового шока HSP25 в GFAP+/Krox20+-шванновских клетках, мигрировавших в белое вещество области повреждения. Отсутствие HSP25+/GFAP+/Krox20+-клеток в области вхождения задних корешков и их наличие в белом веществе может свидетельствовать о возможном становлении реактивности шванновских клеток после их миграции в область демиелинизации, либо о приобретении ими дополнительных защитных свойств при изменении микроокружения. Увеличение экспрессии специфического белка астроцитарной глии S100 в случае прямой генной терапии и при введении трансфицированных терапевтическими генами мононуклеарных клеток крови пуповины может оказывать положительное влияние на регенерацию спинного мозга. Более выраженное увеличение количества S100+-клеток при клеточноопосредованной терапии, по сравнению с прямой инъекцией плазмиды, может быть связано с известной способностью мононуклеарных клеток крови пуповины сдерживать воспалительную реакцию и оказывать нейротрофическое влияние (Kim et al. 2010) или с выработкой специфических факторов, поддерживающих выживание и дифференцировку астроцитов. С использованием иммуногистохимического окрашивания антителами к PDGFpR показано увеличение интенсивности свечения преимущественно в белом веществе в клетках, связанных с кровеносными сосудами (Рис. 72).
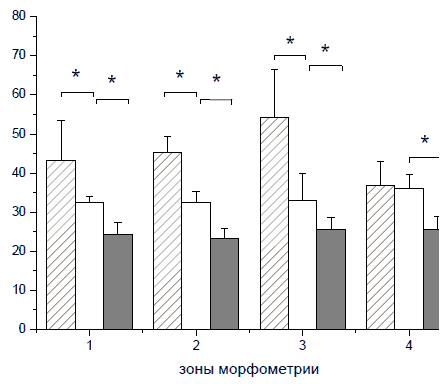
Рис. 72. Экспрессия Р-рецептора тромбоцитарного фактора роста (PDGFpR) в вентральных рогах спинного мозга на расстоянии 1.5 см в каудальном направлении от эпицентра травмы в группе животных с прямой инъекцией плазмиды. Панель А, зеленое окрашивание - PDGFpR+-клетки, панель Б, синее окрашивание - ядра, панель В - объединение А и Б. Конфокальная микроскопия. Увеличение 40х10 Длинные PDGFpR-иммунопозитивные отростки этих периваскулярных клеток ориентированы по ходу кровеносных сосудов, проходящих в радиальном направлении. Наибольшее количестство PDGFpR+-клеток выявлено в наружных зонах белого вещества (Рис. 73).
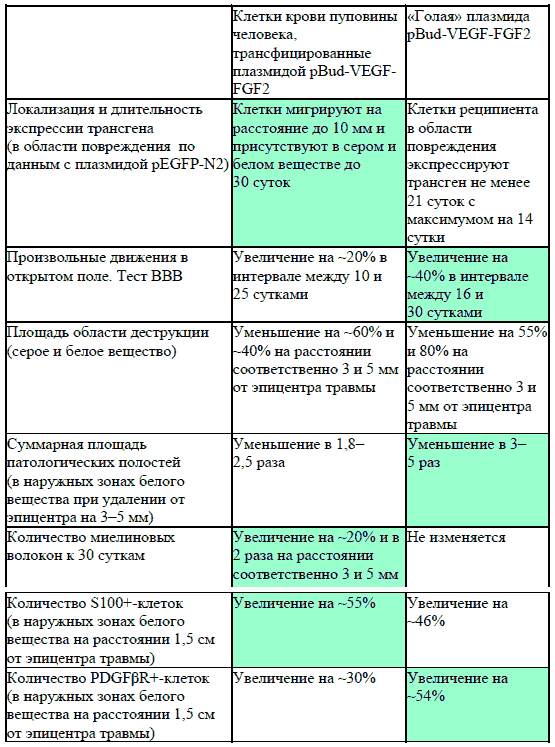
Рис. 73. Количество PDGFpR-иммунопозитивных клеток в белом веществе спинного мозга крысы (по оси ординат) на расстоянии 1.5 см от эпицентра травмы на 30-е сутки эксперимента. Серые столбики - при прямом введении плазмиды pBud-VEGF-FGF2, светлые столбики - при трансплантации ККП, трансфицированных плазмидой pBud-VEGF-FGF2, темные столбики - при трансплантации ККП, трансфицированных плазмидой pEGFP. По оси абсцисс - зоны морфометрии. * - P < 0.05 Различий в характере распределения и интенсивности осадка иммуногистохимической реакции в материале, полученном от животных опытной (pBud-VEGF-FGF2) и контрольной (pBud- EGFP) группы, выявлено не было. К 30-м суткам эксперимента в наружных зонах белого вещества на расстоянии 1.5 см от эпицентра травмы зарегистрировано превышение показателя количества периваскулярных клеток, экспрессирующих PDGFpR, в опыте с прямой доставкой терапевтических генов в среднем на 24% по отношению к группе с трансплантацией трансфицированных клеток (Рис. 73). Этот эффект, возможно, обусловлен действием VEGF, продукция которого оказывается более активной в случае прямой генной терапии. PDGF-B связываясь с рецепторами PDGFpR, стимулирует дифференцировку перицитов из предшественников, экспрессирующих PDGFpR, что оказывает поддерживающее влияние на процесс васкуляризации (Betsholtz et al. 2005). Его нарушение показано при дефектах генов PDGF-B и PDGFpR (Lindahl et al. 1997). Зарегистрированное нами к 30м суткам эксперимента увеличение количества PDGFpR+-клеток отражает усиление васкуляризации белого вещества в области травматического повреждения спинного мозга. Этот эффект, по-видимому, является результатом действия VEGF, продуцируемого трансплантируемыми клетками крови пуповины в межклеточную среду. Появление безмиелиновых волокон в белом веществе в области повреждения у животных с доставкой генов vegf и fgf2 при помощи ККП в качестве носителей трансгенов и при прямой инъекции данной плазмиды свидетельствует о позитивном влиянии данной комбинации генов на процесс нейрорегенерации. Более высокие показатели по таким критериям, как сохранность ткани, васкуляризация и степень восстановления двигательной функции в экспериментах с прямой инъекцией плазмиды pBud-VEGF-FGF2, согласуются с указаниями на аутокринный механизм действия VEGF в регуляции выживания нейронов (Herrera 2009, 2010). Для удобства сопоставления в Табл. 4 суммированы результаты, полученные при прямом и клеточно-опосредованном введении плазмиды с генами VEGF и FGF2. Таблица 4. Эффективность опосредованной ККП человека и прямой доставки «Голой» плазмиды pBud-VEGF-FGF2 с терапевтическими генами факторов VEGF и FGF2 в область контузионного повреждения спинного мозга крысы
Результаты нашего исследования дают основание полагать, что прямая генная терапия представляется эффективным дополнением к методу доставки генов при помощи трансфицированных стволовых и прогениторных клеток. Тем не менее, усилить влияние прямой генной терапии представляется возможным с помощью создания безопасных невирусных векторов с высокой трансфекционной активностью. Назад | Оглавление | Вперед Дата публикации (обновления): 15 февраля 2017 г. 20:44 . |
|
|